|
|
| |
|
|
| |
| |

|
■研究代表者プロフィール
岡部 勝 Masaru
Okabe (PhD)
大阪大学微生物研究所遺伝情報実験センター遺伝子機能解析学分野・教授
1973年大阪大学薬学部修士課程修了。薬学博士。1974年大阪大学薬学部助手に。 1988年 微生物病研究所
助教授を経て1998年より遺伝情報実験施設(センター)教授。2002年より微生物病研究所感染動物実験施設長を併任。受精のメカニズムを主に研究し受精研究に必要な技術を転用した遺伝子改変動物の作製を行う。GFPを全身から発現する“green
mouse”は再生・移植医療などの基礎研究に広く使用されている。
|
研究代表者:
|
岡部勝(微研・遺伝情報実験センター遺伝子機能解析学・教授) |
| |
|
| 研究分担者: |
伊川正人(微研・感染動物実験施設・准教授)
小林 慎(微研・遺伝情報実験センター・COE特任助教)
蓮輪英穀(微研・遺伝情報実験センター・助教)
井上直和(微研・遺伝情報実験センター・助教)
磯谷綾子(微研・遺伝情報実験センター・微研・感染動物実験施設・助教) |
|

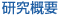
現在、先進諸国では夫婦の10組に1組は不妊に悩まされていると言われており、実際にわが国でも新生児の100人に1人以上の割合で体外受精児が誕生している。少子化の是正が叫ばれる今、不妊を克服することは重要な課題であり、その根本的解決のためには受精のメカニズムを明らかにしなければならない。
受精の分子メカニズムのなかでも謎が多い「融合」は、雌性生殖器内に射出された多くの精子(ヒトの場合1〜3億匹)のなかで、過酷な生存競争に生き残ったごく僅かな精子が正確かつ確実におこさなければならない現象であり、次世代へ種を保存するために必要不可欠なものである。そこには種を超えて共通する見事な仕組みが存在しているのではないかと考えられる。しかし、この仕組みについて分子生物学的な解析はほとんどなされていないのが現状である。そのような背景の中、遺伝子改変技術の台頭により、受精の様々なステップにおける必須因子が次々と明らかにされてきている(図-a)。
我々はこれまでに融合のステップを特異的に阻害する抗精子モノクローナル抗体(OBF13)を用いて、その抗原である「Izumo」の同定に成功している(縁結びの神様が祀られている出雲大社に因んで名付けられた融合因子)。Izumoは細胞外に免疫グロブリン様ドメインをもつ、免疫グロブリンスーパーファミリーの膜貫通型糖タンパク質であり(図-b)、融合可能な精子の膜表面にはじめて現れることが分かった(図-c)。
Izumoノックアウトマウスは雄性不妊であり、その精子を用いて体外受精を行っても受精卵を得ることはできなかった。さらに詳細に観察すると、Izumoノックアウト精子は卵子側の融合因子であるCD9のノックアウト卵子の表現型のように(図-d)、融合できない多数の精子が透明帯と卵子の間の囲卵腔に観察された(図-e)。これらの精子は正常に先体反応を引き起こし卵子細胞膜と結合し、融合のステップを顕微授精法でバイパスさせるとノックアウト精子からも産仔を得られることから、Izumoは融合の過程にのみ必要な精子側の真の融合因子であることが明らかになった。またヒトのIzumoも、卵子との結合または融合に機能するタンパク質であるということを、特異的抗体を用いた体外受精阻害実験から明らかにすることができた。
Izumoを含め受精に関連する多くの細胞表面因子は糖修飾によってコントロールされている。例えばレクチン型の分子シャペロンであるカルメジンは、精子形成過程の段階でN結合型糖鎖を通して精子と卵子の相互認識にかかわるADAM 1, 2, 3やその他の因子の品質管理を行っている。一方、卵子を囲んでいる透明帯の構成成分である糖タンパク質のZP3やZP2は糖鎖を介して、受精可能な精子のみを透明帯に結合させていることが知られている。また受精に至るまでの精子・子宮卵管接合部の相互認識にも糖鎖が媒介していることが報告されている。このことから受精における相互認識は糖鎖が担っているといっても過言ではない。
今後の我々の大きなテーマの一つは遺伝子改変動物を用いた解析により受精における糖鎖の役割を明らかにするとともに、その結果によってもたらされる不妊の診断や治療に役立てていけたらと考えている。
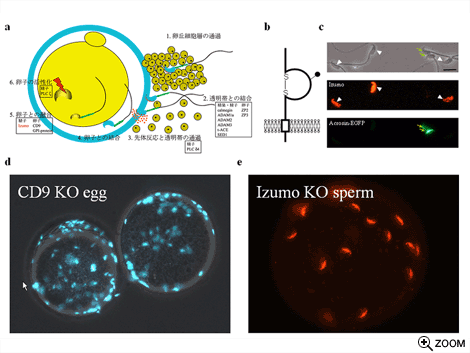
Izumoは世界ではじめて同定された精子膜上の融合因子である。この因子は免疫グロブリンスーパーファミリーに属する糖タンパク質であり、先体反応前の融合できない精子(頭部がグリーンの精子)の表面には現れないが、融合可能な先体反応後の精子の膜表面上に存在する。通常受精卵が観察される条件でIzumoノックアウト精子を卵子に供すると、卵子側の融合因子CD9のノックアウト卵子のように、卵子と融合できないため透明帯と卵子の間の囲卵腔にIzumoノックアウト精子が多数存在する。Izumoは多くの免疫グロブリンスーパーファミリーのタンパク質と同様に、他の分子と結合し複合体を形成することによって細胞間の接着や融合に機能している可能性がある。またIzumoの免疫グロブリン様ドメインにはN型糖鎖が存在しており、その機能について現在解析中である。

|
| 1) |
Okada Y, Ueshin Y, Isotani A, Saito-Fujita T, Nakashima H, Kimura K, Mizoguchi A, Oh-Hora M, Mori Y, Ogata M, Oshima RG, Okabe M, and Ikawa M. Complementation of placental defects and embryonic lethality by trophoblast-specific Ientiviral gene transfer. Nat. Biotechnol., 25, 2, 233-237, 2007.
|
| 2) |
Yamaguchi R, Yamagata K, Ikawa M, Moss SB, and Okabe M. Aberrant distribution of ADAM3 in sperm from both angiotensin-converting enzyme (Ace)- and calmegin (Clgn)-deficient mice. Biol. Reprod., 75, 5, 760-766, 2006. |
| 3) |
Kobayashi S, Isotani A, Mise N, Yamamoto M, Fujihara Y, Kaseda K, Nakanishi T, Ikawa M, Hamada H, Abe K, and Okabe M. Comparison of gene expression in male and female mouse blastocysts revealed imprinting of the X-linked gene, Rhox5/Pem, at preimplantation stages. Curr. Biol., 16, 2, 166-172, 2006. |
| 4) |
Isotani A, Nananishi T, Kobayashi S, Lee J, Chuma S, Nakatsuji N, Ishino F, and Okabe M. Genomic imprinting of XX spermatogonia and XX oocytes recovered from XX<-->XY chimeric testes. Proc. Natl. Acad. Sci. USA., 102, 4039-4044, 2005. |
| 5) |
Inoue N, Ikawa M, Isotani A, and Okabe M. The immunoglobulin superfamily protein Izumo is required for sperm to fuse with eggs. Nature, 434, 7030, 234-238, 2005. |
|
|
|
|
|
| PAGE TOP |
Copyright 2003 Osaka University. All Right Reserved.
Integrated functional analyses of disease-associated sugar chains and proteins, Osaka University

|
|
|
|
|


